Difference between revisions of "BioThalamus"
From aHuman Wiki
(Automated page entry using MWPush.pl) |
(Automated page entry using MWPush.pl) |
||
| (One intermediate revision by the same user not shown) | |||
| Line 29: | Line 29: | ||
** reticular cells | ** reticular cells | ||
** interneurons | ** interneurons | ||
| − | ** feedback projection from layer 6 of cortex | + | ** feedback projection from layer 6 of cortex |
** ascending projection from various scattered cell groups in the brainstem reticular formation | ** ascending projection from various scattered cell groups in the brainstem reticular formation | ||
* relayed inputs | * relayed inputs | ||
| Line 60: | Line 60: | ||
== Divisions == | == Divisions == | ||
| − | * anterior nucleus (AN) - '''association''' - connections similar to the LD nucleus | + | * anterior nucleus (AN) - '''association''' - connections similar to the LD nucleus |
| − | * lateral subnuclei | + | * lateral subnuclei |
** reticular thalamic nucleus - '''nonspecific''' - brain stem reticular formation, cerebral cortex, thalamus -> inhibitory input to thalamic nuclei (arousal and alertness) | ** reticular thalamic nucleus - '''nonspecific''' - brain stem reticular formation, cerebral cortex, thalamus -> inhibitory input to thalamic nuclei (arousal and alertness) | ||
** ventral tiers subnuclei (total 15 nuclei, project to neocortex) | ** ventral tiers subnuclei (total 15 nuclei, project to neocortex) | ||
| Line 91: | Line 91: | ||
*** neurons respond like other neurons to depolarization and hyperpolarization | *** neurons respond like other neurons to depolarization and hyperpolarization | ||
** "burst mode" | ** "burst mode" | ||
| − | *** oscillatory mode" | + | *** oscillatory mode" |
*** neurons in this state have an intrinsic rythmicity | *** neurons in this state have an intrinsic rythmicity | ||
*** during sleep, most thalamic neurons are in burst mode | *** during sleep, most thalamic neurons are in burst mode | ||
| Line 117: | Line 117: | ||
== Axons terminated in Thalamus == | == Axons terminated in Thalamus == | ||
| − | + | ||
* 2 types - R (round) and E (extended), excitatory, using GLU | * 2 types - R (round) and E (extended), excitatory, using GLU | ||
| − | * R-type terminals are characteristically large (3 nm in diameter), although variable in size and actual shape. They conform to the classical type-2 endings, as described in | + | * R-type terminals are characteristically large (3 nm in diameter), although variable in size and actual shape. They conform to the classical type-2 endings, as described in specific thalamic nuclei. The associated axonal terminations are concentrated in sharply delimited, round arbors and '''carry of the order of 100 terminals''', that typically '''end on proximal dendrites''' |
| − | * E-type axons have stalked or spinous terminations of classic type-1 corticothalamic endings. Their axonal terminal | + | * E-type axons have stalked or spinous terminations of classic type-1 corticothalamic endings. Their axonal terminal fields are elongated and quite extended (1-3 mm) and '''carry between 500 and 1,000 E terminals''' that typically '''end on distal dendrites''' |
* in the LGN (and in pulvinar), the driving input from the retina is provided by R-type axon terminals, with type-2 synapses; the input back from cortical area V1 has E-type axon terminals, with type-1 synapses - modulating input, (though there are many more E-type than R-type axons) | * in the LGN (and in pulvinar), the driving input from the retina is provided by R-type axon terminals, with type-2 synapses; the input back from cortical area V1 has E-type axon terminals, with type-1 synapses - modulating input, (though there are many more E-type than R-type axons) | ||
* cortical E-type axons derive from medium to small pyramidal cells in the lower cortical layers. They are located in layer 6, and as a rule always have collaterals in the thalamic reticular nucleus | * cortical E-type axons derive from medium to small pyramidal cells in the lower cortical layers. They are located in layer 6, and as a rule always have collaterals in the thalamic reticular nucleus | ||
| Line 128: | Line 128: | ||
* projections from thalamus to cortex also fall into two classes | * projections from thalamus to cortex also fall into two classes | ||
| − | * | + | * first type goes mainly into layer 4 or lower layer 3, with a minority also contacting processes in layer 6 |
** projection cells in magno- and parvocellular laminae of LGN are prominent examples of such a connection that can very reliably drive cortical cells, despite their small number of synapses. | ** projection cells in magno- and parvocellular laminae of LGN are prominent examples of such a connection that can very reliably drive cortical cells, despite their small number of synapses. | ||
** 2.8% of all excitatory synapses on a layer 4C spiny stellate cell originate from magnocellular cells in LGN | ** 2.8% of all excitatory synapses on a layer 4C spiny stellate cell originate from magnocellular cells in LGN | ||
* other type projects to layer 1, but not exclusively - modulating connection | * other type projects to layer 1, but not exclusively - modulating connection | ||
| − | ** examples - cells in the interlaminar zones of the LGN that project into the | + | ** examples - cells in the interlaminar zones of the LGN that project into the superficial layers of V1 |
* rules | * rules | ||
** (1) If a cortical area projects to a thalamic region from cortical layer 6, then if there is a reverse projection, it goes mainly into layer 4 or lower layer 3 | ** (1) If a cortical area projects to a thalamic region from cortical layer 6, then if there is a reverse projection, it goes mainly into layer 4 or lower layer 3 | ||
** (2) if a cortical area projects to a thalamic region from cortical layer 5, then if there is a reverse projection it avoids layer 4 and often goes mainly to cortical layer 1. These thalamocortical projections are usually much more diffuse than the layer 4 projection. | ** (2) if a cortical area projects to a thalamic region from cortical layer 5, then if there is a reverse projection it avoids layer 4 and often goes mainly to cortical layer 1. These thalamocortical projections are usually much more diffuse than the layer 4 projection. | ||
Latest revision as of 09:14, 22 June 2015
Thalamus facts
Home -> BiologicalLifeResearch -> BiologicalHierarchyFull -> bioThalamus
- this page is about dorsal thalamus (DTH=TH/D)
- other parts of thalalus are:
- ventral thalamus (TH/V) = RN, SVG, ZI
- epithalamus = habenula, pineal gland
see also:
Contents
Overview
- thalamus sorts sensory input, helps sort REALITY from FICTION
Structure


inputs
- 90%-95% of synaptic inputs onto the relay cells, arise from:
- local GABAergic neurons
- reticular cells
- interneurons
- feedback projection from layer 6 of cortex
- ascending projection from various scattered cell groups in the brainstem reticular formation
- relayed inputs
- sensory inputs
- modulatory inputs
- from cerebral cortex
- limbic pathways make input
- cerebellar and basal ganglia inputs
- from reticular thalamic nucleus
- from various brain stem areas
functions:
- site where sensory inputs can be modulated
- relay for cerebellar and basal ganglia inputs to the cerebral cortex
- these are feedback pathways, since the cerebellum and basal ganglia respond to outputs from the cerebral cortex
paths
- thalamus nucleus -> cortex -> thalamus nucleus (the same)
- filtering thalamic inputs to the cerebral cortex
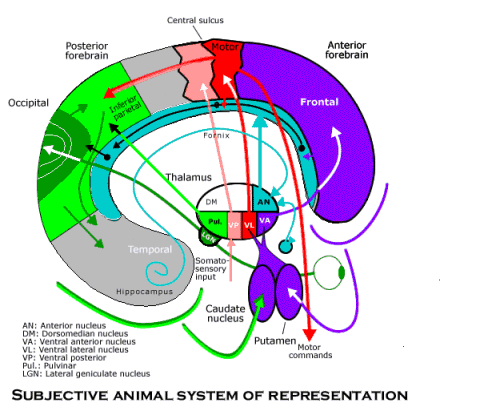

contents
- many inhibitory interneurons
- many neuromodulatory neurotransmitter systems (such as 5HT and NE systems) have terminations within thalamic nuclei
- the relay cell to interneuron ratio is between 3 and 4 to one
Divisions
- anterior nucleus (AN) - association - connections similar to the LD nucleus
- lateral subnuclei
- reticular thalamic nucleus - nonspecific - brain stem reticular formation, cerebral cortex, thalamus -> inhibitory input to thalamic nuclei (arousal and alertness)
- ventral tiers subnuclei (total 15 nuclei, project to neocortex)
- ventral posterior nuclei (VP)
- ventral posteromedial nuclei (VPM) - ff/relay - trigeminothalamic -> cortex
- ventral posterolateral nuclei (VPL) - ff/relay - medial lemniscal and spinothalamic connections -> cortex
- ventral lateral nuclei (VL) - fb/relay - cerebellum/dentate nucleus, basal ganglia -> primary motor, premotor cortex (motor feedback from the cerebellum and basal ganglia to the cerebral cortex)
- ventral anterior nuclei (VA) - fb/relay - basal ganglia (medial globus pallidus, substantia nigra, parts reticulata) -> premotor cortex, supplementary motor area
- ventral posterior nuclei (VP)
- dorsal tiers subnuclei
- pulvinar (PV) - association - superior colliculus, association cortex -> secondary visual areas, association areas in parietotemporal region (visual perception and eye movements, probably relating to attention)
- lateral posterior nuclei (LP) - association - like pulvinar
- lateral dorsal nuclei (LD) - association - hippocampus -> mamillary bodies -> LD -> posterior cingulate cortex (emotional learning)
- medial subnuclei
- medial dorsal nucleus (MD)
- medial subdivision - association - solitary nucleus, substantia nigra reticulata, amygdala and ventral pallidum -> insular cortex, orbital frontal cortex and subcallosal region (autonomic regulation and emotions)
- lateral subdivision - association - superior colliculus, olfactory cortex and the ventral pallidum -> frontal eye fields, anterior cingulate cortex (controlling eye movements, attending to visual stimuli, emotional tone)
- midline nuclei - nonspecific
- intralaminar nuclei
- central median - fb/relay - (reciprocal connections with the globus pallidus and with the premotor cortex)
- parafascicular nuclei - nonspecific
- medial dorsal nucleus (MD)
- metathalamus (near pulvinar)
- MGB - medial geniculate body (auditory relay nucleus) - ff/relay - tonotopically auditory afferents from inferior colliculus -> primary auditory cortex
- LGB - lateral geniculate body (principal visual relay) - ff/relay - retinotopic input -> primary visual cortex
Projection
- each thalamic projection neuron can exist in one of two basic physiological states:
- "tonic mode"
- neurons respond like other neurons to depolarization and hyperpolarization
- "burst mode"
- oscillatory mode"
- neurons in this state have an intrinsic rythmicity
- during sleep, most thalamic neurons are in burst mode
- neurons cannot communicate specific information
- if a novel stimulus is presented, the sudden change from burst to tonic mode may be a major factor in alerting the cortex
- "tonic mode"
Functional View
Sensory Relay:
- sensor/retina -> DTH/V/LGB -> PCA/V1 (1-order visual relay)
- sensor/inferior colliculus -> DTH/V/MGB -> PCA/A1,2 (1-order auditory relay)
- BSA/medial lemniscus, ALS, TTT, STT -> DTH/V/VP -> {PCA/S1,2,3 (1-order somatic relay); HCA/insula (1-order taste relay); ACA/M/4 (?)}
- {BSA/anterior olfactory nucleus; SCA (pain)} -> DTH/M/MD -> {HCA/insula (1-order olfactory relay); ACA/PFC (1-order pain relay)}
- {SCA; BSA/olfactory} -> DTH/I/sheet -> (diffuse)
- BSA/SN,SC,PAG,CR -> DTH/V/VM -> ACA,PCA/layer1 (attention)
Motor relay:
- {BSA/CR; BGA/GP,SN} -> DTH/V/VL,VA -> ACA/M,PM,SM
- BGA/GP,SN -> DTH/I/CM -> ACA/M
Association:
- (many) -> DTH/L/PV -> {PCA/occipital,parietal; HCA/temporal}
- limbic/mammillary -> DTH/A/AV,AM,AD -> ACA/CG
- limbic -> DTH/L/LD -> ACA/CG
Axons terminated in Thalamus
- 2 types - R (round) and E (extended), excitatory, using GLU
- R-type terminals are characteristically large (3 nm in diameter), although variable in size and actual shape. They conform to the classical type-2 endings, as described in specific thalamic nuclei. The associated axonal terminations are concentrated in sharply delimited, round arbors and carry of the order of 100 terminals, that typically end on proximal dendrites
- E-type axons have stalked or spinous terminations of classic type-1 corticothalamic endings. Their axonal terminal fields are elongated and quite extended (1-3 mm) and carry between 500 and 1,000 E terminals that typically end on distal dendrites
- in the LGN (and in pulvinar), the driving input from the retina is provided by R-type axon terminals, with type-2 synapses; the input back from cortical area V1 has E-type axon terminals, with type-1 synapses - modulating input, (though there are many more E-type than R-type axons)
- cortical E-type axons derive from medium to small pyramidal cells in the lower cortical layers. They are located in layer 6, and as a rule always have collaterals in the thalamic reticular nucleus
- cortical R-type axons originate from pyramidal cells in cortical layer 5
Axons terminated in Cortex
- projections from thalamus to cortex also fall into two classes
- first type goes mainly into layer 4 or lower layer 3, with a minority also contacting processes in layer 6
- projection cells in magno- and parvocellular laminae of LGN are prominent examples of such a connection that can very reliably drive cortical cells, despite their small number of synapses.
- 2.8% of all excitatory synapses on a layer 4C spiny stellate cell originate from magnocellular cells in LGN
- other type projects to layer 1, but not exclusively - modulating connection
- examples - cells in the interlaminar zones of the LGN that project into the superficial layers of V1
- rules
- (1) If a cortical area projects to a thalamic region from cortical layer 6, then if there is a reverse projection, it goes mainly into layer 4 or lower layer 3
- (2) if a cortical area projects to a thalamic region from cortical layer 5, then if there is a reverse projection it avoids layer 4 and often goes mainly to cortical layer 1. These thalamocortical projections are usually much more diffuse than the layer 4 projection.